


Next: Acknowledgments
Up: Resonant spatio-temporal learning in
Previous: Learning and recognition
Discussion
We have presented a model where dynamical encoding and processing
can not be reduced to a simple feed-forward process. Our system is
recurrent, and consequently presents an operational closure
[36], so that the inner constraints dominate the
external signal. One can also say that the inner dynamics
corresponds to a ``simulation'' of the external world, which is
updated according to the input signal. In any way, the traditional
input-output dependency is modified: the predictability of a
sensory-motor scheme is dependent on the fitting between the inner
dynamics and the input-output flow. When such fitting is observed
(dynamical or structural coupling, resonance), one can predict the
behavior of the whole system as in traditional input-output
systems. On the contrary, when the fitting is bad, the whole
system is found to produce complex inner dynamics and
unpredictable sensory-motor behaviors.
Even if the link with biology is a delicate matter with our simple
analog neuronal units, we will try in this part to exhibit some plausible
analogies with real biological neural structures and functions, and also to
estimate the limits of such analogy.
The question of recognition is central in our system, and
corresponds to ``acceptance'' or ``rejection'' of some
spatio-temporal configurations arriving on the primary layers.
Most of the signals are rejected, or ignored, while some of them
allow the activation of a specific resonant feedback signal. This
property relates to the question of dynamical binding
[37,4], i.e. the ability to link together
separate elements. In our model, the individual elements composing
a sequence are not significant by themselves. The activation of a
feedback signal needs a specific spatio-temporal disposition of
these individual elements, which are thus processed and
``perceived'' as a whole. This hypothesis of dynamical binding is
present at different scales in neurophysiological studies
[16,23,27]. Our model is one of the possible
implementations of the dynamical binding hypothesis, and thus a
candidate for explaining how such binding occurs in the brain.
The possible role of chaotic dynamics in brain processing is also
a matter of interest for neurophysiologists. The principal
hypothesis on the functional role of chaos in olfactory
perception systems was stated by [32]. In their
article, they interpret the recognition of an olfactory stimulus
as the stabilization of an unstable orbit of a chaotic attractor.
Chaos is thus seen as a reservoir of cycles, where every unstable
orbit corresponds to the encoding of a specific odor. Our model is
not fully compatible with Freeman's model, even if the ideas are
globally comparable. In particular, as the input dynamics takes
part in the dynamical process, the different attractors associated
to every different stimulus do not correspond to a subpart of a
global chaotic attractor. In our model, due to structural
instability, there is no global attractor, but a lot of
transitions between distinct attractors, chaotic or not, depending
on the input signal, on the inner constraints and on previous
learning.
It is difficult to claim high links between our model and
some specific cortical or sub-cortical regions.
However, the 3-populations ReST model, which is devoted to
navigation, has connections with the hippocampus architecture.
Other works by the ETIS team have already taken inspiration from
the hippocampal architecture for the design of control
architecture for the Koala mobile robot
[26,13]. The ReST model has
been inserted in this architecture and takes the place of the area
denoted as CA3. Structurally, this area is known for its important
recurrent links. Functionally, (i) it has been found to display a
functional remapping in case of environmental contextual changes
[3] and (ii) it is supposed to
be the place where locations and/or temporal sequences are
learned when the animal is performing an action. These
observations and hypotheses are compatible with the dynamical
organization and computational abilities of our system. This
analogy has to be more deeply invested, but may already give new
insights in the way the CA3 region processes information.
In the same way, the analogy with the visual system (V1 and V2) is
not straightforward, but our model may however give clues for
people who are interested in the role of feedback links in visual
perception. First, it is known that 80 % of the LGN entries
correspond to feedback connections from the visual cortex.
According to [7], this feedback information is supposed
to enhance the sensitivity of some neurons, according to the
expectations of the cortex on the visual flow. Second, some recent
studies [21] have shown that V1 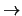 V2
feedback connections may play a role in the process of
figure/ground segregation. Some theoretical models have yet been
proposed, see [28], for explaining the psychological
phenomena of illusionary contour detection. Our model, even
coarser, brings the idea that feedback signal may also be implied
in the reinforcement, by anticipation, of the primary treatment of
dynamical scenes and objects.
At last, one can ask the question of the temporal scales of the
phenomenons we want to model.
This question is not trivial.
Our system uses its own discrete-time parallel updating,
which is not associated to a particular temporal unit.
The time scale is not defined a priori, so that one have to
consider the specific field of application to determine an
``external'' temporal reference.
The visual and perceptual analogies suggest for instance to interpret
synchronization and dynamical binding at the millisecond scale.
On the contrary, sensory-motor experiments take place in the range of seconds.
In future works, and with the same global structures, it may be
necessary to distinguish between the modeling of sensory perception,
with the use of spiking neurons with biologically-compatible temporal scales,
and control models that may correspond to the modeling of global
structures, at broader temporal scales.
Nevertheless, the compatibility between these two interpretations
illustrates in some
way the genericity and ``universality'' of our model of
dynamical perception.
V2
feedback connections may play a role in the process of
figure/ground segregation. Some theoretical models have yet been
proposed, see [28], for explaining the psychological
phenomena of illusionary contour detection. Our model, even
coarser, brings the idea that feedback signal may also be implied
in the reinforcement, by anticipation, of the primary treatment of
dynamical scenes and objects.
At last, one can ask the question of the temporal scales of the
phenomenons we want to model.
This question is not trivial.
Our system uses its own discrete-time parallel updating,
which is not associated to a particular temporal unit.
The time scale is not defined a priori, so that one have to
consider the specific field of application to determine an
``external'' temporal reference.
The visual and perceptual analogies suggest for instance to interpret
synchronization and dynamical binding at the millisecond scale.
On the contrary, sensory-motor experiments take place in the range of seconds.
In future works, and with the same global structures, it may be
necessary to distinguish between the modeling of sensory perception,
with the use of spiking neurons with biologically-compatible temporal scales,
and control models that may correspond to the modeling of global
structures, at broader temporal scales.
Nevertheless, the compatibility between these two interpretations
illustrates in some
way the genericity and ``universality'' of our model of
dynamical perception.
Subsections
Next: Acknowledgments
Up: Resonant spatio-temporal learning in
Previous: Learning and recognition
Dauce Emmanuel
2003-04-08