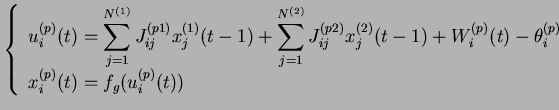Certaines hypothèses neurophysiologiques prêtent aux comportements rythmiques et synchronisés un rôle fondamental en termes de traitement de l'information dans le cerveau.
Ces comportements semblent apparaître lorsqu'une région corticale à la fois reconnaît une configuration d'activation afférente, et produit du fait de cette reconnaissance une configuration d'activation qui peut se propager à d'autres régions corticales, proches ou éloignées.
Par son adaptation dynamique, le système global est capable de traiter de façon différenciée différentes modalités de l'information entrante (voir page ![]() ).
).
En particulier, si l'on accepte l'idée que les oscillations synchrones
servent à augmenter la précision de la réponse du système, la capacité à
reconnaître un motif pourrait reposer sur la capacité à passer d'un régime de
type chaotique à un régime oscillant, dont la régularité est plus grande,
comme cela a été suggéré par Freeman [32] (voir aussi page ![]() ).
).
Il peut être intéressant d'estimer dans quelle mesure un comportement d'adaptation dynamique peut être observé de façon spontanée dans nos système (sans recours à l'apprentissage).
Quelques analogies peuvent en premier lieu être établies avec les structures biologiques :
De nombreux modèles connexionnistes tendent à reproduire ces comportements de synchronisation [11]. Ce sont des modèles à spikes, qui reproduisent plus fidèlement le comportement des neurones réels. Le comportement de notre modèle montre que de tels phénomènes de synchronisation peuvent aussi apparaître sur des systèmes beaucoup plus simples, qui n'ont pas été spécifiquement conçus pour produire des dynamiques synchronisées.
Il est nécessaire, pour mettre en évidence des comportements d'adaptation à un motif d'entrée, de définir ce qu'est un tel motif.
Dans notre système, on considère que l'apparition d'un motif en entrée correspond à une modulation sur les seuils des neurones de la population excitatrice.
Les motifs ![]() utilisés sont des motifs aléatoires statiques comportant
utilisés sont des motifs aléatoires statiques comportant ![]() valeurs tirés selon la loi gaussienne
valeurs tirés selon la loi gaussienne
![]() . Le vecteur
. Le vecteur  est un vecteur nul.
est un vecteur nul.
Un système sur lequel aucun motif n'est présenté est dit spontané, et évolue selon l'équation (2.5).
Un système qui possède un motif ![]() en entrée est dit contraint, et évolue selon l'équation (2.9).
On peut noter que pour nos simulations, la valeur de
en entrée est dit contraint, et évolue selon l'équation (2.9).
On peut noter que pour nos simulations, la valeur de ![]() est systématiquement mise à 0.
Pour
est systématiquement mise à 0.
Pour
![]() et
et
![]() ,
,
Le motif d'entrée constitue donc un nouveau seuil sur la population excitatrice.
Pour les paramètres
(![]() ,
,
![]() ,
, ![]() ,
,
![]() ,
,
![]() ,
,
![]() ,
,
![]() ),
la dynamique spontanée est un point fixe.
Sur la population excitatrice, chaque signal d'activation individuel tend vers une
valeur fixe
),
la dynamique spontanée est un point fixe.
Sur la population excitatrice, chaque signal d'activation individuel tend vers une
valeur fixe
![]() , proche de zéro.
Lorsqu'on envoie un motif statique aléatoire
, proche de zéro.
Lorsqu'on envoie un motif statique aléatoire ![]() (avec
(avec
![]() ) sur la population excitatrice, cela tend à donner naissance à un régime dynamique.
La naissance de ce régime dynamique correspond exactement à l'apparition d'une sous population, au sein de la population excitatrice, dont le signal
de potentiel atteint la partie linéaire de la fonction de transfert,
ce qui se traduit par des pics sur le signal d'activation
- voir la figure 2.26 -.
Chaque neurone de cette sous-population tend à produire un signal d'activation de forte amplitude.
On dit que le neurone est activé par le motif.
Cette sous-population représente de 5 à 10%
de la population excitatrice totale.
L'ensemble de ces neurones activés constitue la réponse spatiale du réseau.
Un autre motif
) sur la population excitatrice, cela tend à donner naissance à un régime dynamique.
La naissance de ce régime dynamique correspond exactement à l'apparition d'une sous population, au sein de la population excitatrice, dont le signal
de potentiel atteint la partie linéaire de la fonction de transfert,
ce qui se traduit par des pics sur le signal d'activation
- voir la figure 2.26 -.
Chaque neurone de cette sous-population tend à produire un signal d'activation de forte amplitude.
On dit que le neurone est activé par le motif.
Cette sous-population représente de 5 à 10%
de la population excitatrice totale.
L'ensemble de ces neurones activés constitue la réponse spatiale du réseau.
Un autre motif
![]() indépendant de
indépendant de ![]() tend à produire une réponse spatiale différente (les neurones activés par le deuxième motif ne sont pas les mêmes).
Le système est apte à produire un signal spécifique pour chaque entrée différente.
On peut noter par ailleurs que l'ensemble des neurones activés ne coïncide pas avec les valeurs maximales du motif d'entrée.
Le réseau produit donc une forme de codage de son entrée.
tend à produire une réponse spatiale différente (les neurones activés par le deuxième motif ne sont pas les mêmes).
Le système est apte à produire un signal spécifique pour chaque entrée différente.
On peut noter par ailleurs que l'ensemble des neurones activés ne coïncide pas avec les valeurs maximales du motif d'entrée.
Le réseau produit donc une forme de codage de son entrée.
![\includegraphics[]{nn_reac_theta1.eps}](img651.png)
![\includegraphics[]{nn_reac_theta2.eps}](img652.png)
|
Enfin, le régime dynamique n'est pas nécessairement le même d'un motif à l'autre : il s'opère une autre forme de discrimination sur les motifs d'entrée qui porte cette fois-ci sur la nature du régime dynamique produit.
La nature du régime dynamique donne la réponse dynamique du réseau.
Cette réponse dynamique s'avère très sensible aux valeurs précises du motif en entrée.
De petits changements sur ce motif peuvent en effet conduire à des changements
drastiques dans la réponse dynamique du système (alors que dans le même temps la réponse
spatiale est à peine changée)
Ainsi, une discrimination fine entre deux motifs voisins peut être effectuée
par de tels changements dans la dynamique.
C'est ce qu'on peut voir sur la figure 2.27, où deux motifs fortement
corrélés (cor(![]() ,
,
![]() ) = 0.98) mènent à une réponse spatiale presque
identique (les mêmes neurones sont activés) tandis que les les régimes dynamiques produits sont différents, le premier étant un régime chaotique cyclostationnaire, et le second un régime d'oscillations synchronisées.
) = 0.98) mènent à une réponse spatiale presque
identique (les mêmes neurones sont activés) tandis que les les régimes dynamiques produits sont différents, le premier étant un régime chaotique cyclostationnaire, et le second un régime d'oscillations synchronisées.
En étendant ces résultats aux systèmes neuronaux biologiques, on peut penser que la capacité à distinguer deux motifs voisins pourrait reposer sur un ajustement fin des régimes dynamiques. Par exemple, atteindre un régime synchronisé oscillant pourrait indiquer que le motif correspond bien à la forme mémorisée, tandis qu'un régime cyclostationnaire indiquerait seulement que l'on n'est pas trop loin de la forme mémorisée. Ainsi, au sein d'un système sensoriel, seule une bonne correspondance entre la forme mémorisée et la forme observée pourrait produire le signal régulier nécessaire un traitement ultérieur de cette information. Ce mécanisme pourrait être relié à des mécanismes naturels, comme ceux qui ont été décrits par [23] dans le cadre du système olfactif.
![\includegraphics[]{nn_reac_theta1a.eps}](img653.png)
![\includegraphics[]{nn_reac_theta1b.eps}](img654.png)
|